Gass, M, Rebar, R, Glob. libr. women's med.,
(ISSN: 1756-2228) 2008; DOI 10.3843/GLOWM.10078
January 2008
Molecular and Mendelian Disorders
Authors
INTRODUCTION
In the early 1980s, the field of genetics involved pedigree analysis, risk assessment, and rare circumstances in which a precise diagnosis could be made by chromosomal analysis or biochemical studies. Now, genetics is a rapidly expanding specialty in which direct analysis of gene defects is possible, and the potential for correcting these defects is on the horizon. With this exponential expansion of knowledge regarding the molecular basis for disease have come novel patterns of inheritance that challenge the basic principles of inheritance that have been taught for years. This chapter presents the basic principles of Mendelian inheritance, the molecular basis for a number of common genetic disorders, and current methods of molecular diagnosis.
PRINCIPLES OF MENDELIAN INHERITANCE
Mendelian disorders result from a mutation at a single genetic locus. A locus may be present on an autosome or on a sex chromosome, and it may be manifest in a dominant or a recessive mode. There are more than 10,000 traits believed to be inherited in a Mendelian fashion,1 but only a few of the more common disorders of interest to the obstetrician–gynecologist are highlighted in this chapter.
The patterns of inheritance for the various Mendelian traits are illustrated by the idealized pedigrees in Figure 1. An autosomal recessive trait (disease) is expressed only when the mutant gene is present in a double dose (homozygous state). Both parents are heterozygous and possess one copy of the mutant gene and one copy of the normal or functional gene. Autosomal recessive traits are characterized as follows: (1) there is rarely a positive family history outside the affected sibship; (2) males and females are equally likely to be affected; and (3) heterozygous parents have a 25% chance of producing an offspring affected with the disease.
{kind=link}
Autosomal dominant traits manifest in the heterozygous state (single-gene dose) and are characterized by the following: (1) they can be transmitted from generation to generation; (2) the probability that a person carrying the gene will pass it on to offspring is 50%; and (3) males and females are equally likely to be affected.
Males (XY) and females (XX) differ in the number of X chromosomes they possess. As a result, the inheritance pattern of mutations carried on the X chromosome will differ from the inheritance pattern of mutations on autosomes. A recessive trait controlled by a gene on the X chromosome will be expressed in all males carrying the allele. Affected males are said to be hemizygous. Females will be affected if they are homozygous or if they inactivate most of the X chromosomes carrying the normal allele.2 The following are characteristics of X-linked inheritance: (1) there is no male-to-male transmission; (2) all daughters of an affected male receive the mutant gene and are therefore carriers; and (3) one-half the sons and daughters of a heterozygous female receive the mutant gene.
The distinction between X-linked dominant and X-linked recessive is unclear, but in general, X-linked recessive refers to a trait that is not clinically expressed in the heterozygous female, and X-linked dominant to a trait that is expressed in the heterozygous female.
LABORATORY METHODS
Before the introduction of molecular diagnostic capabilities, prenatal diagnosis was limited largely to disorders for which the primary genetic defect was known and a known gene product could be assayed. Most commonly, the diagnosable conditions consisted of metabolic disorders that resulted from a deficiency of an enzyme that could be assayed in cultured amniotic fluid cells. However, because the DNA complement is identical in every cell of an organism, a hereditary defect detectable at the DNA level should be present in any nucleated cell at any stage of differentiation. Diagnosis is possible even for inherited disorders with an unknown primary pathologic process.
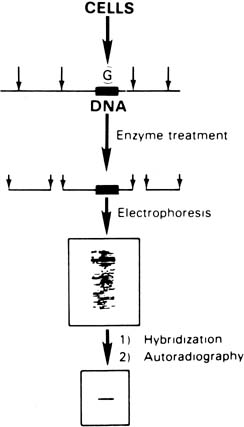
There are certain basic tools required for DNA analysis. The first tools are restriction endonucleases. These bacterial enzymes recognize and cleave a specific DNA base sequence. Direct identification of mutant genes is possible by virtue of the specificity of these enzymes. A single nucleotide change in an enzyme recognition site would be readily detectable, because the enzyme would no longer recognize the nucleotide sequence as a cut site. Therefore, there would be an alteration in the length of the specific DNA fragments normally produced by digestion with a particular enzyme (Fig. 2). After restriction endonuclease digestion, agarose gel electrophoresis is used to separate DNA fragments by size (Fig. 3). The Southern blot technique is used to transfer the DNA from the gel to a nitrocellulose filter, and specific fragments can be located by hybridization with radiolabeled strands of known DNA sequences. Because they hybridize specifically to complementary sequences within the DNA fragments on the filter, these strands can be used as probes. After autoradiography, those fragments containing sequences that hybridize with the probe can be seen as bands on the developed film. By comparing their location with molecular weight markers of known length seen in the same gel, one can determine the size of the fragments. Length is expressed in kilobases, or thousands of base pairs.
{kind=link}
{kind=link}
DNA probes likewise are essential tools and can be derived in a number of ways. Genomic probes are isolated from human DNA and, by analysis, have been shown to contain a specific gene or to be located close to a specific gene. Complementary DNA (cDNA) probes are prepared by isolation of known messenger ribonucleic acid (mRNA) and copying it back into a specific DNA. For many genes, the exact nucleotide sequence is known, and it is possible to manufacture synthetic oligonucleotide probes that are complementary to these sequences. Their short length of 20–50 nucleotides prevents them from hybridizing with any sequence that varies even as little as a single nucleotide. Therefore, for any portion of a gene or a mutation involving that gene, a synthetic oligonucleotide probe can theoretically be made if a short sequence of the gene is known.
Polymerase chain reaction
The development of the polymerase chain reaction technique has greatly simplified DNA analysis and shortened laboratory time.3 Unlike the Southern blot technique, which is used in an attempt to identify one gene from the many genes using DNA probes, PCR allows the exponential amplification of the target gene, providing a direct analysis of the gene in question. PCR does require, however, that the sequence of the gene, or at least the borders of the region to be amplified, be known.
To perform PCR (Fig. 4), a double-stranded piece of DNA is heated to separate it into two single strands (a process called denaturing). Added to the mixture of denatured DNA are two oligonucleotide primers (each 20–30 nucleotides long). They are exactly complementary to the ends of each piece of double-stranded DNA to be amplified. Each primer has a 3-prime free end that serves as a substrate (starting point) for DNA synthesis using the initial DNA strand as a template. Because the primers are added in vast excess, when the temperature is reduced the original single strands anneal with the primers rather than with each other. In the final step, DNA polymerase, a highly heat-stable bacterial enzyme that synthesizes DNA, is added along with precursor deoxyribonucleotides. DNA synthesis proceeds down each DNA strand, making a new copy of the DNA, resulting in two double-stranded DNA molecules. In cycle two, the two double strands are denatured into four single strands, and the process ultimately results in four double strands. The process is repeated for 30 cycles, resulting in nearly one billion copies of the original DNA sequence.
{kind=link}
PCR has greatly facilitated DNA diagnosis by markedly shortening the time necessary for a diagnosis and by making it possible to form a diagnosis on the basis of a sample as small as a single cell. Because it is such a sensitive method, however, extreme caution must be exercised to prevent contamination by foreign DNA, such as sloughed skin cells from ungloved technicians.
Allele-specific oligonucleotide hybridization
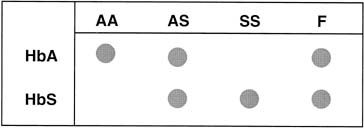
Single base change mutations can be detected in genes if the sequence around the gene of interest is known. To detect these mutations, oligonucleotide primers that differ by a single nucleotide are synthesized in vitro; one is complementary to the normal allele and the other is complementary to the mutant allele base sequence. The normal oligonucleotide sequence will hybridize to the normal allele, but not to the mutant allele. The mutant oligonucleotide sequence will hybridize only to a complementary mutant allele, but not to the normal allele. Figure 5 depicts the use of allele-specific oligonucleotide hybridization (ASOH) for the diagnosis of sickle cell anemia. DNA is amplified by PCR and spotted onto a nitrocellulose membrane along with control samples for normal AA, heterozygous AS, and homozygous SS. One membrane is hybridized with the normal oligonucleotide and one with the mutant oligonucleotide. The sample in this prenatal diagnosis case has signal with both the HbA and HbS probes, indicating a sickle cell carrier. To utilize ASOH, the exact sequence of both the normal and the mutant gene must be known.
{kind=link}
Short tandem repeat polymorphism
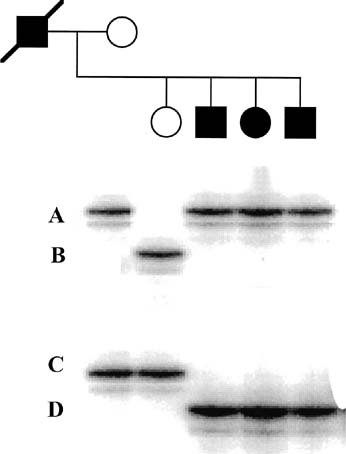
Short tandem repeats (STR) are stretches of DNA composed almost entirely of repeating cytosine adenosine (CA) dinucleotides. These stretches of repeats are found throughout the human genome; the number of repeats at any locus may vary, and this is known as a polymorphism. The exact number of repeats at any locus can be measured with PCR amplification. For diagnostic testing, DNA primers complementary to the sequences that flank the STR of interest are synthesized. STR analysis can be performed via radiolabeled nucleotides. Incorporation of these deoxynucleotides yields a radioactive PCR product. The PCR is electrophoresed, and the gel is then used to expose photographic film. Such an analysis is shown in Figure 6. This family is segregating an autosomal-dominant disease. Even without DNA from the deceased father, the disease haplotype can be straightforwardly determined. This type of analysis is very useful for determining affected status in families with late-onset or reduced penetrance disease.
{kind=link}
Affected persons are designated by shaded symbols. From this result, it can be concluded that the father's haplotype was B/D and that the disease is associated with marker D.
Alternatively, during PCR synthesis, the PCR products are labeled with a fluorescent marker that allows detection on an automated sequencer. This technology allows for multiplex PCR, the concomitant analysis of several STR (see section on Duchenne's muscular dystrophy).
Single-strand conformational polymorphism analysis
Single-strand conformational polymorphism (SSCP) analysis of PCR-amplified DNA is based on the tendency of single-stranded DNA to undergo intramolecular base-pairing. The electrophoretic mobility of the DNA molecule in a gel system is a function of its conformation as well as its length. If a point mutation affects intramolecular base-pairing, it will alter its conformation and thus its electrophoretic mobility. Two single-stranded DNA molecules that differ by even a single base can be differentiated by SSCP analysis. A major advantage of SSCP analysis is that it can be performed without knowing any specific mutation in the gene to be analyzed. For this analysis, DNA from a suspected mutant gene and DNA from a normal gene are amplified via PCR (Fig. 7). The double-stranded DNA is heat-denatured to form single strands that are then rapidly cooled. As cooling occurs, the single-stranded DNA will form intramolecular base pairs before it can bind with a complementary strand. Because the strands, although complementary, have very different sequences, each allele yields two conformers when applied to a gel. Both the normal and the mutant DNA will undergo base pairing, but if the mutation affects normal pairing, the pattern of folding will be different for each strand, resulting in an alteration in the electrophoretic mobility of the mutant gene. As such, persons heterozygous for a mutation show four bands on a gel, whereas normal and mutant homozygotes show two. Any detected mutation may or may not actually cause disease. Many single nucleotide changes do not alter the sequence of the protein product. Additionally, as is the case for linkage analysis, these polymorphisms can be within the gene or adjacent to the gene. Therefore precise testing using SSCP analysis requires that the altered DNA conformers be analyzed to determine whether the base change is responsible for the disease in question.
{kind=link}
DIAGNOSIS OF SPECIFIC MENDELIAN DISORDERS
Autosomal dominant conditions
SKELETAL DYSPLASIA
Until recently, the diagnosis of skeletal dysplasias in children was dependent on radiographic and physical features. Because of the often-subtle variations between categories and the limitations of ultrasonography, prenatal detection was limited to only the most severe types. With the discovery of the molecular basis for many of these conditions, a precise prenatal diagnosis can now be determined as early as the first trimester via chorionic villus sampling (CVS).
ACHONDROPLASIA
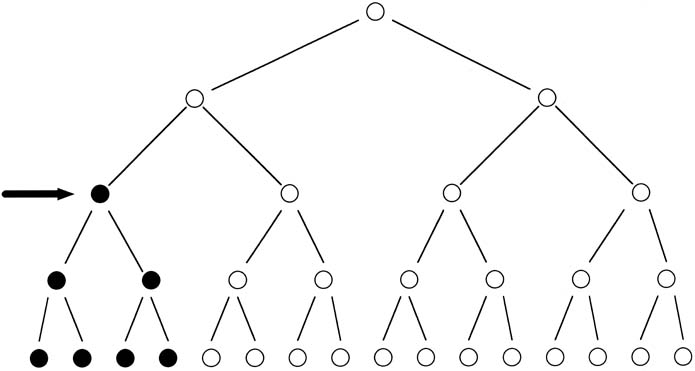
Achondroplasia is the single most common skeletal dysplasia in humans. Inherited in an autosomal-dominant fashion, a person with achondroplasia has a 50% chance of having a child with this disorder. Because the gene is 100% penetrant, a person will have the disease if the gene is inherited. Parents of normal height who have a child with achondroplasia would normally be expected to have a nearly 0% chance of having a second child similarly affected; however, the risk is actually 5–6% because one parent (usually the father) has the mutation in more than one germ cell, known as germ cell mosaicism (Fig. 8). Achondroplasia results from a mutation in a gene known as fibroblast growth factor receptor 3 (FGFR-3). Identification of a mutation in this gene enables a precise diagnosis in subsequent pregnancies, either by amniocentesis or by CVS.4, 5 Moreover, the evaluation is simplified by the fact that all mutations to date have been point mutations, and almost all of the mutations have involved the same amino acid substitution.
{kind=link}
FGFR-3 mutations also have been found to cause lethal dwarfism known as thanatophoric dysplasia.6 Like achondroplasia, it represents a new dominant mutation with an empiric risk for recurrence of approximately 5%. As important, however, is the ability to use molecular testing to confirm the diagnosis in cases in which severe short-limb dwarfism is diagnosed on the basis of ultrasound examination. Using amniotic fluid cells, one can make a precise diagnosis and therefore provide the family with more exact prognostic information.
OSTEOGENESIS IMPERFECTA
Osteogenesis imperfecta (OI) describes a constellation of disorders characterized by brittle bones and defects in type I collagen. The form of OI of significance for the obstetrician–gynecologist is the perinatal lethal form, OI type II. Also a new dominant mutation, it can most easily be diagnosed by assays of collagen structure.7 If these assays are performed on fibroblasts from the affected child, prenatal diagnosis is straightforward in any subsequent pregnancy. There is, however, one major caveat: because this analysis is of protein structure, not DNA sequence, future pregnancies must be evaluated by CVS, not amniocentesis. Collagen defects can be studied in chorionic villi, but not in aminocytes.
NEUROLOGIC DISEASES
The onset of many of the autosomal-dominant neurologic diseases occurs in adulthood, and these diseases are generally more familiar to the neurologist than to the obstetrician–gynecologist. However, two of these disorders, myotonic dystrophy and Huntington disease, are discussed to illustrate the necessity of having a basic knowledge of the mechanisms of inheritance and the clinical implications of these disorders.
Myotonic dystrophy (MD) is a condition characterized by myotonia, cataracts, and other variable features, such as male-pattern baldness. The onset of symptoms usually occurs in the third or fourth decade of life. In addition to the adult form, there is a neonatal form of the disorder known as congenital myotonic dystrophy. This disorder is characterized by severe hypotonia, respiratory compromise, and, often, death in the newborn period. Those infants who do survive commonly have severe developmental delay.
The MD gene is characterized by a repeated sequence of cytosine-thymidine-guanine (CTG), which in normal persons is repeated between 5 and 30 times. This "trinucleotide repeat" is expanded to greater than 100 repeats in persons affected with adult-onset MD.8 Neonates with congenital MD may have more than 2000 copies of the repeat. Figure 9 presents a pedigree of a family that would typically present to the obstetrician–gynecologist. The patient (proband) presents at 10 weeks' gestation with a history that her sister has adult-onset MD. Neither of her parents, both in their late fifties, have any symptoms to suggest that either is affected with MD. Based on the absence of any symptoms in the patient or her parents, we might assume that the sister's disease represents a new dominant mutation. Molecular testing of the family, however, indicates a much different scenario. The sister has 200 repeats, consistent with her clinical symptoms. Her mother has 50 repeats, which is in the range of what is called a premutation (35–100 repeats). The proband also has 200 repeats and can be expected to experience MD symptoms in the future. Molecular evaluation reveals that the fetus has 2000 repeats, consistent with the diagnosis of congenital MD.
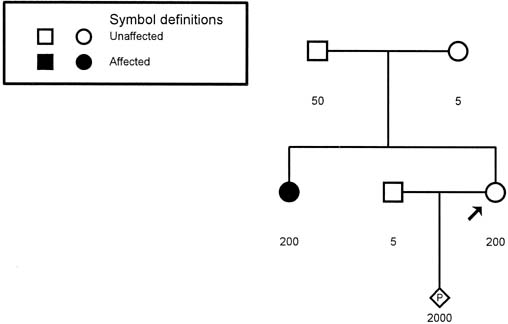{kind=link}
Trinucleotide repeat disorders are characterized by the presence of unstable premutations that may remain unchanged or may amplify during spermatogenesis or oogenesis. This process of expansion of the trinucleotide repeat is known as anticipation. In the case of MD, marked amplification can occur when the gene is passed from the mother, but not from the father. Therefore, the congenital form of MD is seen only when the gene is passed from the mother to the fetus. Determination of the status of a person with a family history of MD (i.e., normal, premutation, full mutation) is quite precise with the use of modern molecular techniques.
Huntington disease (HD) is a late-onset, progressive, fatal disease inherited in a classic autosomal-dominant fashion. The early symptoms of the disease are subtle loss of muscle coordination, forgetfulness, and personality changes. The disease progresses in stages, from choreiform movements (hence the older name of Huntington's chorea) to hypokinesis and then to rigidity. Ultimately, the patient is bedridden with dysphagia, dysarthria, and impairment of gait and coordination. Onset of the disease occurs most commonly between the ages of 30 and 50 years.9
HD is also caused by a mutation involving a trinucleotide repeat sequence. Normal persons have between 11 and 31 copies of a CAG repeat. The full mutation range is between 38 and 100 copies. There is an intermediate range of 32–38 repeats, and there are examples of both affected and unaffected persons with this number of repeats.10 Therefore, there does not appear to be a true premutation in HD. Unlike MD, inheritance of HD from the father is associated with expansion of the repeats and an earlier age of onset. In approximately one third of cases in which the father passes on the gene to his offspring, there is an expansion resulting in juvenile-onset HD.11
The ability to diagnose HD precisely by molecular techniques offers the possibility of presymptomatic (predictive) testing and prenatal detection of an affected fetus. Although predictive testing can offer freedom from the psychological burdens associated with being at risk if the person does not carry the mutation, the impact on persons found to have the gene can be devastating. Those found to carry the gene face the inevitability of a disease for which there is currently no treatment.
More commonly the obstetrician–gynecologist will be presented with the request for prenatal testing by an at-risk patient. It must be emphasized to the couple that testing will determine precisely whether the fetus has the HD gene. Thus, a positive result gives an at-risk patient a presymptomatic diagnosis of HD. Therefore direct prenatal testing should not be performed unless an at-risk patient has already undergone predictive testing.
An alternative to prenatal testing is available for at-risk couples who do not wish to undergo predictive testing. This method is known as exclusion testing.12 The HD gene is located on chromosome 4, and DNA markers (restriction fragment length polymorphism [RFLP]) are available that allow chromosome 4 to be traced through generations. By testing the grandparents of the fetus, it can be determined whether the fetus received a chromosome 4 from the affected grandparent or the unaffected grandparent. Because the chromosome 4 received from the affected grandparent could carry either the HD gene or the normal gene, the fetus has the same risk as the parent for having HD (i.e., 50%). If the fetus receives a chromosome 4 from the unaffected grandparent, the risk for having HD is 0%, and the patients can be assured that the gene will not be passed on to the next generation.
Because of the significant ethical, psychological, and medical issues associated with predictive testing, such testing should take place only at institutions where protocols are in place for pretesting and posttesting counseling and support.
Autosomal recessive disorders
CYSTIC FIBROSIS
This autosomal recessive condition is the most common recessive condition affecting whites, with a carrier frequency of one in 29. Cystic fibrosis is caused by a mutation in the CFTR gene, a chloride ion channel in epithelial cells. Mutations in this gene cause severe lung disease and pancreatic insufficiency in the classic form but may result in milder phenotypes, including only congenital bilateral absence of the vas deferens in some affected males. The most common mutation is delta-F508, the deletion of a phenylalanine residue at position 508, which accounts for 70% of the mutations in the white population. To date, however, there are more than 1000 known mutations throughout the gene that can cause disease. Because of the large number of known mutations in the gene, and because the phenotype is variable, prenatal testing may be problematic as complete gene sequencing is expensive, cumbersome, and impractical. Screening for the most common mutations in a population is more practical. Carrier testing for the CF mutation should be offered to individuals with a family history of cystic fibrosis, and to partners of individuals with cystic fibrosis. In 2001 the American College of Obstetricians and Gynecologists (ACOG) set forth the following testing screening guidelines: in addition to couples who have a family history of cystic fibrosis or when one partner is affected by cystic fibrosis, all couples in which one or both of the partners are white (including those of Ashkenazi Jewish descent) and are planning a pregnancy or are currently pregnant be offered carrier screening for cystic fibrosis.13 ACOG recommended that these couples be offered a prenatal screen that includes the 25 most common mutations and any mutation with a population incidence of more than 1%.
Although prenatal identification of a fetus affected with cystic fibrosis gives couples the option of pregnancy termination, cystic fibrosis is not a condition that is currently treatable in utero, nor does prenatal detection of the disease appear to improve outcomes for affected individuals. Rarely, an affected pregnancy may present with ultrasonographic abnormalities such as echogenic bowel, and testing of the parents or the fetus may be offered to confirm this diagnosis.
HEMOGLOBINOPATHIES
Rapid progress has been made in the area of antenatal detection of the hemoglobinopathies, and they serve as the prototype for the available diagnostic methodologies. From initially requiring a fetal blood sample obtained by the relatively high-risk procedures of fetoscopy or placental aspiration, we have moved to methods that require only uncultured amniotic fluid cells, or chorionic villi.
Hemoglobin is a tetrameric protein consisting of four globin chains. In humans, there are six structurally different types of globin chains: alpha (α), beta (β), gamma (γ), delta (δ), epsilon (ε), and zeta (ζ).14 The synthesis of various chains is switched on and off during the process of differentiation and may represent an adaptive process for the developing fetus. The embryonic chains (ε and ζ) are rapidly replaced as the fetus develops. Each of the different hemoglobins found in humans is formed by the combination of two α-chains and two non-α–chains (γ, β, δ).
The messenger RNA (mRNA) for each globin gene is transcribed from a varying number of genes, which may differ among populations. In general, there are four genes controlling α-chain synthesis and two controlling β-chain synthesis. Genes for the α-chains are located on chromosome 16, and those for β, γ, and δ are located on chromosome 11. In the hemoglobinopathies, the specific disorder may be considered recessive, because heterozygous persons have half the genes (either one or two) functioning to make the normal protein and half functioning to make no protein or an abnormal one. Therefore, either the homozygous state or a double heterozygous state (i.e., two different deleterious mutations) can result in a hemoglobinopathy.
Sickle cell disease is caused by a single nucleotide change (AT) in codon six of the b gene. A similar point mutation (GA) in the same codon results in sickle C disease. As discussed, sickle cell disease can result from the homozygous state (SS or CC) or from the double heterozygous state (SC). Sickle S disease is unique in that the mutation modifies the recognition site for two restriction endonucleases, Mst II and DdeI. As outlined in Figure 2, this change results in a change in length of the DNA fragment with the sickle gene. In Figure 10, a Southern blot technique is shown, indicating that it is possible to distinguish sickle trait (both βS and βA), sickle disease (only βS), and normal hemoglobin (only βA) by the size of the fragments produced. However, the sickle C mutation does not alter this specific cut site; therefore, it could not be detected by this specific methodology.
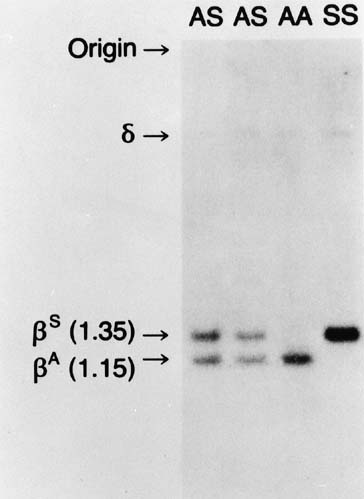{kind=link}
Because the gene sequence for both the S and the C mutation is known, it is possible to develop specific oligonucleotide probes that recognize both the normal and the mutant DNA sequence.15 As depicted in Figure 5, the technique of ASOH can provide a precise diagnosis. DNA is extracted from blood, amniotic fluid cells, or chorionic villi; amplified by PCR; and then dotted onto the filter paper. Oligonucleotides for both the normal gene and the mutant gene are then used in the hybridization. Analysis of the pattern of dots allows precise identification of normal persons, heterozygous carriers, and affected persons.
α-Thalassemia is characterized by a deficiency in α-globin chain synthesis. Normal α-chain production is the product of four functioning a genes (two on each chromosome 16). Therefore, either one or both genes can be deleted on a chromosome 16, resulting in four clinical states according to the number of functional α-globin genes (3, 2, 1, or 0). Of clinical significance is the circumstance in which there is one functional gene, known as hemoglobin H disease, or 0 functional genes, known as hemoglobin Bart's disease, which results in fetal hydrops.
Although a number of mutations have been described in the α-gene, α-thalassemia results from deletion of the α-gene cluster on chromosome 16. At least 14 different deletions have been characterized.16 Because the mutations represent deletions, standard restriction enzyme analysis is the preferred method for both carrier detection and prenatal diagnosis.
Of importance to the obstetrician–gynecologist is the clinical presentation of the homozygous form of the disorder. The mother may present with severe preeclampsia in the early third trimester, and ultrasound evaluation reveals hydrops fetalis (mirror syndrome). Because of the lethal nature of this condition and the potential maternal morbidity, all patients of Asian ancestry should be screened for the carrier state of α-thalassemia. A simple screening tool is a complete blood count with red cell indices. Carriers of the α-thalassemia mutation have a mild anemia and a mean corpuscular volume of less than 80 femtoliters (fL). Currently, the simplest method for the diagnosis of α-thalassemia carriers is to exclude iron deficiency anemia by appropriate studies and to exclude the carrier state for β-thalassemia by hemoglobin electrophoresis. Because both parents must be carriers to have an affected fetus, the next step at this point should be to test the woman's partner. If he also appears to be a carrier, appropriate molecular studies can be performed to confirm the diagnosis in both parents. When both parents are confirmed to be carriers, prenatal diagnosis is possible either by CVS or by amniocentesis.
β-Thalassemia is defined as either reduced (β+) or absent (βO) β-chain synthesis. β-Thalassemia is prevalent in areas of the world where malaria is endemic (Mediterranean area, Africa, Middle East, India, Southeast Asia, and Southern China). Although more than 170 mutations have been characterized,17 only 25 different mutations account for the majority of mutant alleles in at-risk populations.18 Based on knowledge of the mutations that are specific to a geographic region or ethnic group, one can use a PCR-based screening protocol to detect approximately 80% of the common mutations and another 15% of rare mutations.19
As with α-thalassemia, the simplest initial approach to screening for β-thalassemia is to obtain a complete blood count with red cell indices. When the mean corpuscular volume is less than 80 fL, hemoglobin electrophoresis is indicated. Carriers of the β-thalassemia gene will have an elevated level of hemoglobin A2 (>3.5% of total hemoglobin will be A2). If the partner is also found to be a carrier, appropriate molecular studies must then be performed to determine the exact mutations before prenatal diagnosis can be offered. An algorithm for the screening for hemoglobinopathies is depicted in Figure 11.
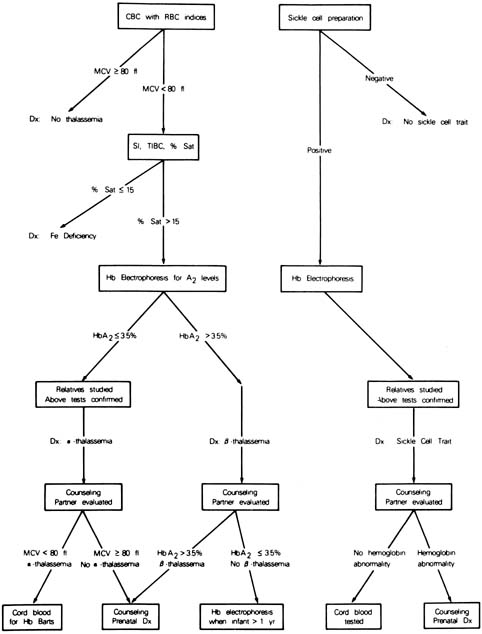{kind=link}
X-linked disorders
DUCHENNE MUSCULAR DYSTROPHY
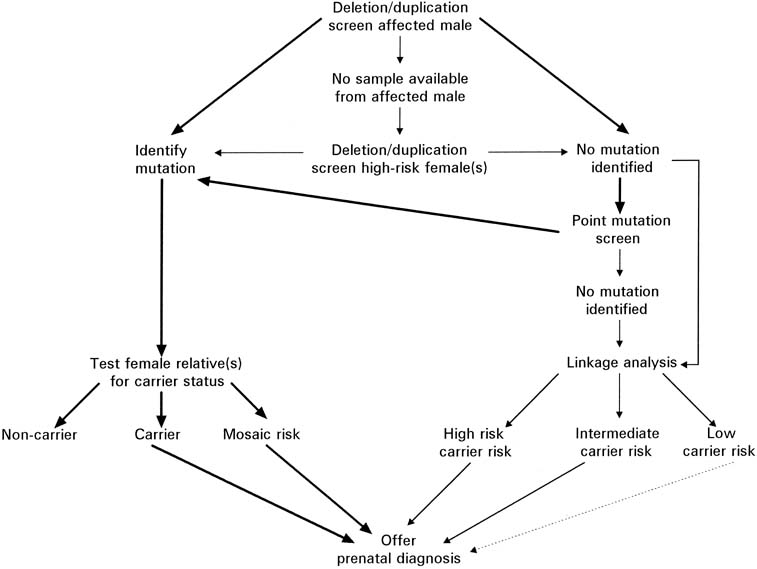
Duchenne muscular dystrophy (DMD) is an X-linked recessive disorder affecting one in 3,500 males. The disease is caused by a mutation in the dystrophin gene, the largest (2.5 megabases) gene known. Deletions within the gene account for 60–65% of the dystrophin mutations.20 An additional 6–10% of mutations are duplications.21 Therefore, using the techniques of multiplex PCR (Fig. 12), one can detect approximately 70% of all mutations. In Figure 12, a multiplex STR polymorphism analysis for the four repeat elements found within the dystrophin gene is shown for a DMD family. In this example, females have two alleles at each locus (X-linked), each allele differing by a few base pairs. This type of analysis is beneficial in determining the carrier status of II-1 and II-3. Affected son II-2 shows a deletion by the absence of a peak at STR-45. From this information, the mother's haplotype can be accurately determined, thus identifying the disease haplotype (a-aa). Inspection of the analysis of II-1 and II-3 shows that they are both DMD carriers. Therefore, during a subsequent pregnancy, DNA derived from either chorionic villi or amniocytes would show whether a son was affected (a-aa) or unaffected (acab). Because the affected males may no longer be alive, determining carrier status may be complicated, and in some cases impossible, in families with a single affected person. An algorithm for evaluation of a potential carrier is outlined in Figure 13. Because of the significant amount of work required before prenatal testing can be offered, it is essential that patients be evaluated whenever possible before conception, if all options for prenatal testing are to be available to them.
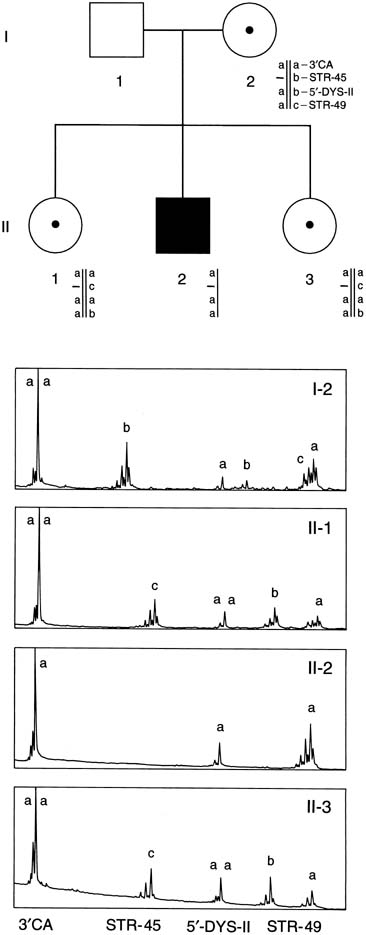{kind=link}
{kind=link}
Despite the availability of these molecular techniques, there remain a number of families in which the fetal risk for DMD remains unresolved. However, in those cases in which the potential for DMD transmission is significant, the alternative is to assay directly for dystrophin, the protein product of the gene, via an immunofluorescent technique; this assay requires a sample of muscle obtained by fetal muscle biopsy.22 Although associated with a higher risk for fetal loss, this approach does offer the possibility of accurate prenatal diagnosis when molecular studies are uninformative.
FRAGILE X SYNDROME
Fragile X syndrome is the most common inherited form of mental retardation, affecting approximately one in 4,000 men and one in 8,000 women.23 Therefore as many as one in 259 women in the general population may be carriers of the gene.24 Fragile X syndrome has a wide range of clinical presentations, including moderate disabilities in females, autism, and other psychiatric disorders.
Unlike classic X-linked disorders (e.g., DMD, hemophilia), fragile X syndrome affects both males and females; it is important to note that there are males who carry the gene but have normal intelligence and no physical stigmata of the disorder. This variable clinical phenotype reflects the novel mutation in the gene known as fragile X mental retardation 1 (FMR1). The FMR1 gene is characterized by a repeated sequence of the trinucleotide cytosine-guanine-guanine (CGG).25 In unaffected persons, between six and 50 repeats of this sequence are found. Intellectually normal carriers of the mutation have between 50 and 200 CGG repeats, called a premutation. In carrier females, premutations are unstable and may undergo further expansion during oogenesis. If the CGG sequence expands beyond 200 repeats, it is considered a full mutation, and all males with this number of repeats will show the clinical features of fragile X syndrome, and approximately one-half of females likewise will have mental retardation.26 Males with the premutation do not have expansion during spermatogenesis but will pass the premutation to all of their daughters, each of whom will be at risk for having an affected child.
Table 1 outlines clinical situations that should prompt screening for fragile X syndrome, either in a pregnant patient or in the patient presenting for preconception counseling. Two of these recommendations are straightforward, but the latter two require explanation. We traditionally have assumed that X-linked disorders in the patient's father's family (paternal side) would not place the pregnancy at risk. In the case of fragile X syndrome, however, the gene is nonpenetrant in 20% of males with the fragile X gene (i.e., they do not show clinical symptoms). Therefore, a patient's father who has a family history of mental retardation may carry the fragile-X premutation, which he will pass to all of his daughters. In the circumstance in which the patient's paternal uncle has unexplained mental retardation, her risk of being a carrier of the fragile X gene is approximately one in 100. Therefore any history of unexplained mental retardation should prompt testing for fragile X carrier status.
Table 1. Patients for whom fragile X screening is indicated
| Women with a known family history of fragile X syndrome |
| Women who have learning disabilities or mental retardation |
| Women with a family history of unexplained mental retardation |
| Women with a family history of premature ovarian failure |
Of particular interest to the obstetrician–gynecologist is the association of fragile X premutation and premature menopause.27, 28 Preliminary studies have suggested that fragile X premutation carriers are three-times more likely than other women to experience premature menopause. Based on this information and the ease of carrier screening, testing would seem appropriate in circumstances in which there is a family history of premature menopause.
Prenatal testing for fragile X syndrome by either amniocentesis or CVS is quite precise because of the consistent mutation present in all families. There are, however, two caveats. The finding of a full mutation in a male fetus is indicative of an affected child. However, the presence of a full mutation in a female fetus gives an empiric risk of approximately 50% that the child will be affected with the clinical features of fragile X syndrome. Although prenatal testing is possible using either CVS or amniocentesis, the difference in methylation patterns of the gene found in early gestation requires that a laboratory providing prenatal testing have experience with the nuances of testing that are associated with CVS.29
FUTURE PROSPECTS
Remarkable advances have been made in prenatal diagnosis and genetic screening, and the future holds great promise for the further development and refinement of techniques for in utero diagnosis. DNA analysis has become a powerful tool for detecting genetic disorders, and it will be extended to other diseases that are identified at the DNA level.
The future of prenatal diagnosis and genetic screening is exciting and holds great promise. An ever-increasing number of disorders will be diagnosed in utero. The advances in all areas of science interface with this field, and their clinical application will greatly expand the number of diagnosable conditions. Of all specialties of medicine to be affected by these advances, obstetrics and gynecology should receive the greatest impact. To identify and counsel patients at risk for a specific disorder in an appropriate manner, the obstetrician must have a basic knowledge of genetics and the techniques of prenatal diagnosis. The practicing obstetrician is the interface between the patient and the rapidly expanding areas of prenatal diagnosis and genetic screening.
ACKNOWLEDGMENT
The authors thank Dr Mitchell S. Golbus for his significant contributions to previous editions of this chapter.
REFERENCES
Online Mendelian Inheritance in Man, OMIM (TM). McKusick-Nathans Institute for Genetic Medicine, Johns Hopkins University (Baltimore, MD) and National Center for Biotechnology Information, National Library of Medicine (Bethesda, MD), 2000. World Wide Web URL:http://www.ncbi.nlm.nih.gov/omim/ |
|
Puck JM, Willard HF: X inactivation in females with X-linked disease (editorial; comment). N Engl J Med 338: 325, 1998 |
|
Erlich A, Gelfand DH, Saiki RK: Specific DNA amplification. Nature 331: 461, 1988 |
|
Shiang R, Thompson LM, Zhu Y-Z et al: Mutations in the transmembrane domain of FGFR-3 cause the most common genetic form of dwarfism, achondroplasia. Cell 78: 335, 1994 |
|
Rousseau F, Bonaventure J, Legeai-Mallet L et al: Mutations in the gene encoding fibroblast growth factor receptor-3 in achondroplasia. Nature 371: 252, 1994 |
|
Tavormina PL, Shiang R, Thompson LM et al: Thanatophoric dysplasia (types I and II) caused by distinct mutations in fibroblast growth factor receptor 3. Nat Genet 9: 321, 1995 |
|
Pepin M, Atkinson M, Starman BJ, Byers PH: Strategies and outcomes of prenatal diagnosis for osteogenesis imperfecta: A review of biochemical and molecular studies completed in 129 pregnancies. Prenat Diagn 17: 559, 1997 |
|
Rodman JB, Fenwick RG, Fu Y-H et al: Relationship between partial trinucleotide repeat length and severity of myotonic dystrophy in offspring. JAMA 269: 1960, 1993 |
|
Quarrell OW, Harper PS: The clinical neurology of Huntington's disease. In Harper PS (ed): (ed): Huntington's Disease. pp 37-80, Philadelphia, WB Saunders, |
|
Huntington's Disease Collaborative Research Group: A novel gene containing a trinucleotide repeat that is unstable and expanded on Huntington's disease chromosomes. Cell 72: 971, 1993 |
|
Crauford D: Huntington's disease. Prenat Diagn 16: 1237, 1996 |
|
Hayden MR, Hewitt J, Kastelein JJ et al: First-trimester prenatal diagnosis for Huntington's disease with DNA probes. Lancet 1: 1284, 1987 |
|
Grody WW, Cutting GR, Klinger KW et al: Laboratory standards and guidelines for population-based cystic fibrosis carrier screening. Genet Med 3: 149–154, 2001 |
|
Huehn ER, Danee D, Beaver GH et al: Human embryonic hemoglobin. Nature 201: 1095, 1964 |
|
Embury SH, Scharf SJ, Saiki RK et al: Rapid prenatal diagnosis of sickle cell anemia by a new method of DNA analysis. N Engl J Med 316: 656, 1987 |
|
Higgs DR, Vickers MA, Wilkie AOM et al: A review of the molecular genetics of the human &b.alpha;-globin gene cluster. Blood 73: 1081, 1989 |
|
Baysal E: The &b.beta; and &b.delta;-thalassemia repository. Hemoglobin 19: 213, 1995 |
|
Huisman THJ: Frequencies of common &b.beta;-thalassemia alleles among different populations: Variability in clinical severity. Br J Haematol 75: 453, 1990 |
|
Varawalla NY, Old JM, Sarkar R et al: The spectrum of &b.beta;-thalassemia mutations on the Indian subcontinent: The basis for prenatal diagnosis. Br J Haematol 78: 242, 1991 |
|
Koenig M, Hoffman EP, Berfelson CJ et al: Complete cloning of the Duchenne muscular (DMD) cDNA and preliminary genomic organization of the DMD gene in normal and affected individuals. Cell 50: 509, 1987 |
|
Abbs S: Prenatal diagnosis of Duchenne and Becker muscular dystrophy. Prenat Diagn 16: 1187, 1996 |
|
Evans MI, Quintero RA, King M et al: Endoscopically assisted ultrasound-guided fetal muscle biopsy. Fetal Diagn Ther 10: 167, 1995 |
|
Turner G, Webb S, Wake S, Robinson H: Prevalence of fragile X-syndrome. Am J Med Genet 64: 196, 1996 |
|
Rousseau F, Rouillard P, Morel M-L et al: Prevalence of carriers of premutation-size alleles of the FMR1 gene—and implications for the population genetics of fragile X syndrome. Am J Hum Genet 57: 1006, 1995 |
|
Oberle I, Rousseau F, Heitz D et al: Instability of a 550-base pair DNA segment and abnormal methylation in fragile X syndrome. Science 252: 1097, 1991 |
|
Finucane B: Should all pregnant women be offered carrier testing for fragile X syndrome. ? Clin Obstet Gynecol 39: 772, 1996 |
|
Schwartz CE, Dean J, Howard-Pebbles PN et al: Obstetrical and gynecological complications in fragile X carriers. Am J Med Genet 51: 400, 1994 |
|
Conway GS, Hettiarachchi Murray A, Jacobs P: Fragile X premutations in familial premature ovarian failure. Lancet 346: 309, 1995 |
|
Sutherland GR, Mulley JC: Fragile X syndrome and fragile XE mental retardation. Prenat Diagn 16: 11199, 1996 |